約32億年前、地球上に光を利用してエネルギーを作り出す生物、すなわち光合成生物が誕生した。それまでにも生物はいたが、現在の生物とはエネルギー代謝の機能が全く異なるので、実質的には光合成生物の誕生をもって生命の誕生として差し支えないだろう。これ以降、長い生物の歴史の中で、進化、分化あるいは退化(退行進化)が繰り返され、現在、500万種あるいは3000万種ともいわれる生物種がこの地球上に生息する。そのうち、植物種はおよそ35万を数えるといわれている。32億年前には単細胞の微生物であり、その間に植物種が増加してきたわけだから、その過程には何らかの連続した進化や分化の道筋があるはずである。この道筋を系統 というが、形態、植物生理の機構あるいは生体内の物質代謝にいたるまである一定の順序で発達をしてきたわけだから、一つの系統に属する種の間には、様々な形で類縁関係があるはずである。生物の進化、分化の道筋に基づいた分類学の手法を系統分類 (systematics)あるいは自然分類 と称する。それに対するものが人為分類 であり、リンネを初めとする古典分類学がそれに相当する。現代の分類学は全て系統に従って生物種を配列した系統分類学である。系統分類学の利点は、系統配列することで形態の類似、生理機能や化学成分の類縁関係などが明らかになり生物種相互の関係を理解しやすくなることである。しかし、進化、分化の道筋が必ずしも現存するとはかぎらず、むしろ途中で消滅してしまった種の方が多い。系統を明らかにするために、化石として残された生物種の研究、すなわち古生物学が有力な実証の手段にはなるが、植物は本質的に化石を残し難いので、今日でも完全に実証づけられた系統の関係はごくわずかなものである。系統分類学が起こったのは19世紀以降だが、それは決して一枚岩の存在ではなく、多くの分類学者によって提唱された種々の分類方式がある。それらの間には系統の考え方にもかなりの差が認められ、現在になっても完全な定説はできていない状態である。植物の外部形態や微細形態に基づいた植物系統分類方式としては、Engler-PlantlとBentham-Hookerとに大別され、現在ではそれぞれを改良したものが用いられている。一方、近年では、分子生物学の進歩に伴い、DNAの塩基配列の決定が比較的簡便となり、一定のマーカー遺伝子(分類学者によってどの遺伝子をマーカーとして用いるかは異なる)の塩基配列を比較することによって分類を行うこと、すなわち分子分類学(molecular phylogeny)も 盛んに行われている。しかし、その解析結果が形態分類を主とする古典分類学と一致しないこともしばしばあり、まだ発展途上の段階にある。
(1)Engler-Prantlの分類方式
ドイツのエングラー(Adolf Engler; 1844-1930)とプランテル(Karl Prantl; 1888-1911)は被子植物に花被の有無と、花葉の離生・合生を重視した分類方式を採用し種子植物の進化の方向を考え、花の部分構造でより進化とした進化型を後にして配列した。後に、新しい知見も取り入れられ、次のような新エングラー方式に発展した。
新エングラー方式では、離弁花植物と合弁花植物を分断しているところにおいて、従来からの人為分類的な考え方を残しているが、細菌類から被子植物までの全植物界をカバーしていること、他の方式とも大きな矛盾が認められないことなどから、現在広く使われている。
(2)Bentham-Hookerの分類方式
イギリスのベンサム(George Bentham; 1800-84)とフッカー(Jpseph Dalton Hooker; 1817-1911)は被子植物の花被は有花被のほうが原始型で、コショウ目などの無花被植物は、花被の退化によって後からできたとし、花被のあるモクレン科、キンポウゲ科が裸子植物から直接進化してきた、被子植物の中でもっとも原始的なものとし、子房の位置と状態を重視した配列を行った。のちにハッチンソン(Hutchinson1884-1954)、ベッシイ(Charles Edwin Bessey;1845-1915)らに引き継がれた。この方式による最新の分類では双子葉植物を次の5群に分けている。
- 花托群
- 無弁花または離弁花;子房上位
- 花冠群
- 合弁花
- 萼筒群
- 無弁花または離弁花;子房中位または下位
- 子房群
- 合弁花
- 尾状花序群
- 単性の無弁花;少なくとも雄花は長い尾状花序となる
この方式はイギリスやアメリカでよく採用された。
(3)クロンキスト分類方式
クロンキスト体系は1981年・1988年にクロンキスト(Arthur Cronquist;1919年―1992年)によって提唱された分類法で、系統進化の配列をかなり大幅に変更しているが、マクロの形態分類による系統進化の仮説に基づく分類という点では新エングラー体系の延長線上にある。『植物の世界』(朝日新聞社、1997年)などの図鑑で採用されたが、広く普及することはなかった。
(4)APG分類方式
1990年代後半になって登場した分類体系で、Angiosperm Phylogeny Groupのイニシャルをとって命名したもの。直訳すると”被子植物系統グループ”となるように、被子植物の分類を対象とした体系であり、今日、もっとも広く植物分類学者から支持を集めている。APG体系はミクロのゲノム解析に基づいて構築した分類体系であり、分類の根拠を仮説に基づく新エングラー・クロンキスト体系と比べて、最新の分子生物学の知見を取り入れたより客観的かつ実証的な分類法ということができる。分子系統を基にした系統樹で表示されるところは一見直感的でわかりやすいが、合弁花・離弁花など花の形態を重視した新エングラー・クロンキスト体系とは大きく配列が異なる。とりわけ科名の変更が著しく、例えばユリ科(Liliaceae)では、ネギ科(Alliaceae;後にヒガンバナ科ネギ亜科に変更された)・クサスギカズラ科(Asparagaceae)・キスゲ科(Hemerocallidaceae)・スズラン科(Ruscaceae)・イヌサフラン科(Colchicaceae)・狭義のユリ科などに細分化されている。そのほか、多くの重要な薬用植物を含む旧ゴマノハグサ科(Scrophulariaceae)も、APG体系では多くの種が別の科に移され、例えば、ジギタリスはオオバコ科 (Plantaginaceae) に編入されるなど、旧来の分類とは大きく変更を余儀なくされている。1998年に初めて提唱され、2003年、2009年に改訂され、それぞれAPG I、APG II、APG IIIと称している。現在の分類学の主流はいうまでもなくAPG体系であり、形態分類を基にしたクロンキスト以前の体系はもはや歴史的体系となりつつある。本サイトの植物はすべてAPG IIIに準拠している。
-オウレン属の系統と種分化について-
前述のいずれの方式を採用するにせよ、現在の分類学では種間の系統を考慮するのが主流となっている。いかなる種もある母となる種があって進化や分化を繰り返してきたはずであるが、前述したように道筋に当たる種は必ずしも現存するとは限らない。とりわけ、上位の分類段階についてはその傾向が著しく実証はきわめて困難である。しかし、下位の分類段階、例えば属以下であれば分化の歴史は比較的最近のことであって分化のレベルは比較的軽微であるので系統を推定する手がかりは現存することが多くなる。ここで、漢方薬原料となるキンポウゲ科オウレンCoptis japonicaの所属するオウレン属を例としてその種分化の系統について説明する。以下の例は挿入図も含めて次の文献より引用したものである。
- 田村道夫編、『日本の植物』、62-83頁(培風館、東京、1981年)
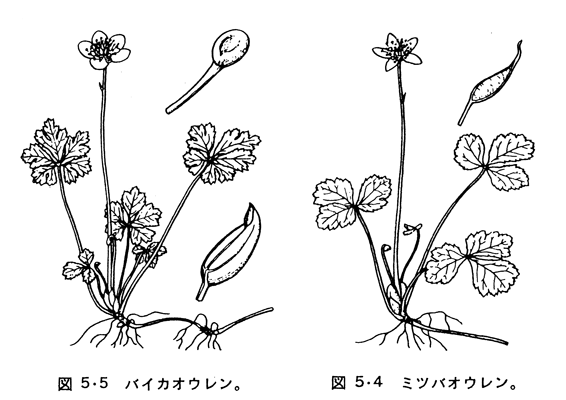
オウレン属は、通例、オウレン節(代表種としてオウレンCoptis japonicaの図を挙げる)、ミツバオウレン節(同ミツバオウレンC. trifolia)、バイカオウレン節(同バイカオウレンC. quinquefolia)の3節に分けられ、それぞれの果実、萼、花片、花序、葉の形態学的特長を次の表に示す。
{kind=link}
{kind=link}
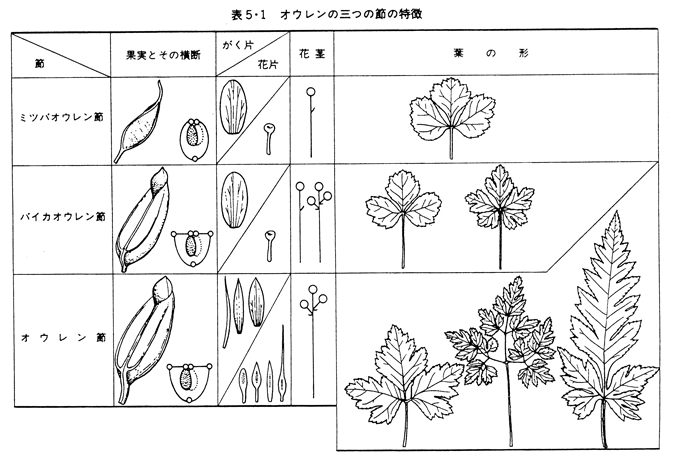
これによると、果実の形態ではバイカオウレン節とオウレン節では裂開果でほとんど差がないのに対して、ミツバオウレン節の果実は閉果で全く異なる。一方、がく片(図で花弁に見えるものは萼である)、花片(花弁に相当するもの)の形態ではミツバオウレン節、バイカオウレン節はほとんど差がないのに対して、オウレン節の形態は多様である。花茎ではバイカオウレン節ではオウレン節、ミツバオウレン節の両タイプのものがあり、葉の形態ではいずれも3出掌状のものが共通するが、オウレン節では2回~3回羽状複葉まで分化したものがある。これらのことから節のレベルでのオウレン属の種分化の経路は次のように考えられる。
- ミツバオウレン節からバイカオウレン節を経てオウレン節へ分化
- オウレン節からバイカオウレン節を経てミツバオウレン節へ分化
- バイカオウレン節から独立にオウレン節、ミツバオウレン節へ分化
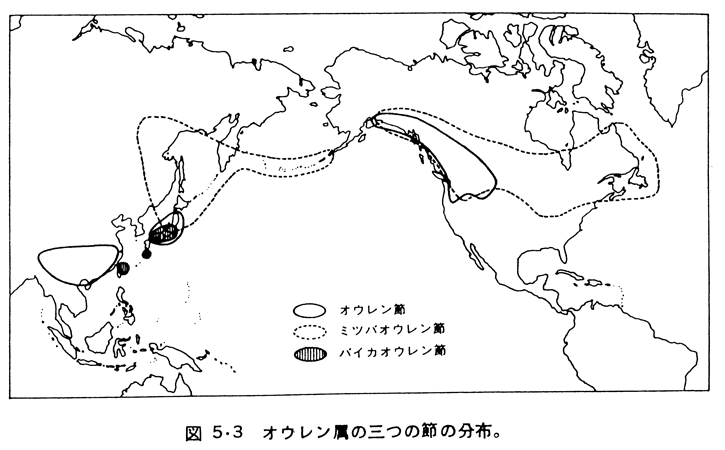
このうちどれが妥当であるかを判断するにはどれがもっとも原始的かを判断する必要がある。本論文の著者はバイカオウレン節が形態学的にもっとも原始的とし、3が妥当であるとしている。種は、その発生以降、分布を広げるとともに新しい環境に適応しながら更に分化する。オウレン属3節の地理的分布を示したのが次の図であるが、南中国から日本列島、アリューシャン列島を経て北米大陸を横断するように分布する。これによれば3節の分布が交錯するのは日本列島だけであり、またオウレン属種がもっとも多く分布するのも日本列島である。一般に、種の発生中心は、周辺の分布域と比べると、より長い時間をかけて分化するため、形質の多様性という観点からもっとも豊かとなる。ロシアの植物学者バビロフ(Nikolai I. Vavilov; 1887 - 1943)はこの視点に立って栽培植物の発生中心を推定し、今日でも有用遺伝資源が多く集積する地域をバビロフセンターと称している。バビロフ説の視点に立てば、オウレン属の発生中心は日本列島となる。したがって、地理的分布状態からバイカオウレン節からオウレン節とミツバオウレン節が分化したという形態学的検討に基づく結論は妥当と思われる。すなわち、バイカオウレン節がまず日本列島で発生し、それからミツバオウレン節、オウレン節が分化し、分布域を東西に広げたと推定できるのである。
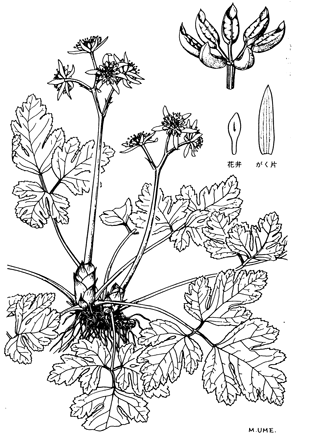
以上、オウレン属の系統について説明した。前述したが、長い地史的スケールで見れば、種は変動するものである。種の範囲がそれぞれの種で異なるのは分化のレベルに差があるからである。オウレンを例に挙げれば、種としてはCoptis japonicaにまとめられているが、次の図に示すように葉の形態は産地によりきわめて多様であり、キクバオウレン、セリバオウレン、コセリバオウレンの3つのタイプが識別されている。これは現在もそれぞれの地域で活発な種分化が進行中であることを示している。オウレンは漢方薬として賞用される高価な生薬原料であり、現在では針葉樹林下での播種により半栽培的に供給されている。しかし、しばしば他地域産の種子が播かれることがあり、結果として交雑による生態系の撹乱を招いている。交雑で発生したものは必ずしも生き残るとは限らず、これにより種の絶滅する危険性があることを留意すべきである。有用遺伝資源保全の観点からかかる撹乱はあってはならないことはいうまでもない。
