高等植物の二次代謝産物は一次代謝産物の代謝経路からスピンアウトして生合成されることは既に述べた(→植物に含まれる化学成分について)。全ての二次代謝産物は酵素によって触媒される化学反応の積み重ねによって生合成され、また同じ前駆体を出発物質とした場合でも化学反応の組み合わせによって様々な構造をもつ二次代謝産物の生合成が可能となる。その化学反応の組み合わせや触媒する酵素は各生物種により全て同じというわけではないので、生物種ごとに生産される二次代謝産物は異なる。すなわち生物界における二次代謝産物の化学的多様性は以上の背景によって成立したものである。生物種は刻々と変わる環境へ適応するため常に連続的に形質の変化が起きて種が分化する。また、突然変異による不連続的な形質の変化も起き、優れた形質のみが生き残って進化する。生物種は分化(differentiation)と進化(evolution)を繰り返して現在の種の多様性が造り上げられたのであるが、二次代謝産物も遺伝子の制御を受ける酵素反応により生合成されるのであるから、二次代謝産物においても分化、進化が起きる。生物種が新たな反応を触媒する酵素を獲得することによって新しい化学種を生み出し、この連鎖により化学進化、分化が進行した結果、現在見るような多種多様な二次代謝産物群が構築されたのである。化学進化の過程で起きる触媒酵素の獲得は新たな遺伝形質として考えられるため、生物種の分化とともに二次代謝産物の化学進化、分化も同時に進行する。二次代謝産物の組成およびその生合成経路を解析すれば系統分類ができるという観点に立って、二次代謝産物の分布と化学系統解析をもとにして生物の系統分類を論ずる学問を化学系統分類学(chemotaxonomy)と称する。しかし、実際には全ての生物種について二次代謝産物の構造、組成が明らかになっている訳ではないので、通例、形態分類学の補助手段として利用することが多く、二次代謝成分の組成だけで分類学的位置を論ずることはない。一般に、科や属以下のレベルにおいて系統的に近縁種であれば類似の二次代謝産物を産する場合が多いのであるが、類似の二次代謝産物を含むからといって必ずしもそれらが近縁とは限らない例も多いことも事実である。したがって、化学系統分類学は比較的狭い範囲の分類を論ずる場合に用いられることが多い。このように生物種の分類そのものを対象とする場合においては化学系統分類学はごく限定的な存在ではあるが、天然界から有用物質を探索する場合には化学分類学の知見は役に立つ。例えば、バイオプロスペクティングにおいて過去に発見された生物活性物質の類縁化合物を探索する場合、系統的に近い分類群を集中的に探索することが常法となっている。二次代謝産物の分布と植物分類がどう相関するか、代表的な二次代謝産物を挙げて説明しよう。
1.ゲッキツ属の系統分類について
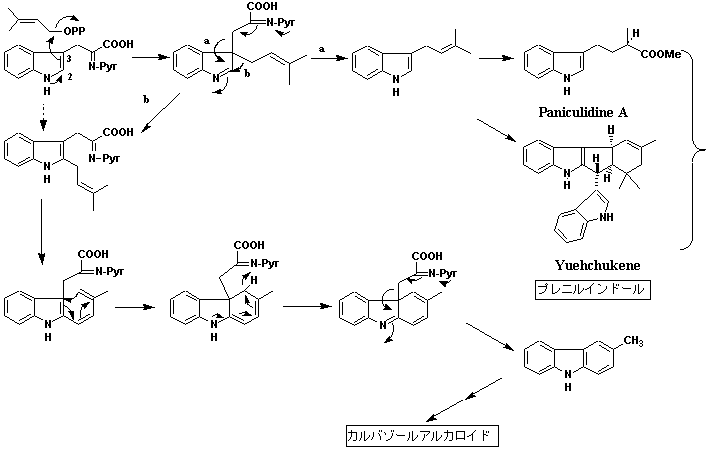
ミカン科ゲッキツ(Murraya)属は形態分類学の観点からオオバゲッキツ節とゲッキツ節に分割される(→ゲッキツ属の分類と分布参照)。これは田中長三郎により提唱されたのであるが、比較的化学的探索が進んだ現在、以下に述べるように化学分類学的観点からも支持される。すなわち、オオバゲッキツ節に属するオオバゲッキツ(別名ナンヨウザンショウ)M. koenigiiをはじめいずれの種もカルバゾールアルカロイドと称される特異な二次代謝産物を含むのに対して、ゲッキツ節各種の主成分はプレニルクマリン誘導体であり、カルバゾールアルカロイドの存在は確認されていない。それに代わってゲッキツ節に属する種からはプレニルインドールと称する特異な二次代謝産物が報告されている。カルバゾールとプレニルインドールはその炭素基本骨格の構造から考えて、いずれもトリプトファンとジメチルアリル二リン酸から生合成されることに疑問の余地はないが、トリプトファンへのプレニル化の位置の違いで両化合物の分化が起きると推定される。すなわち、オオバゲッキツ節に属する種ではインドールの2位(次図の経路a)、ゲッキツ節に属する種では同3位(経路b)で起き、カルバゾール・プレニルインドールに分化すると考えることができる。以上を化学分類学的観点から解釈すれば、カルバゾール・プレニルインドールの生合成過程においてトリプトファンへのプレニル化がインドールの2位、3位のいずれで起きるかが種分化の分岐点となる。一方、系統的にゲッキツ属と近縁とされるワンピ(Clausena)属にはカルバゾールアルカロイドとフロクマリン(オオバゲッキツ節にも知られている)が共存する種が多い。ワンピ属とゲッキツ属はこれまでも近縁とされてきたが、化学分類学的見地からとりわけオオバゲッキツ節との密接な関連が認められる。
南西諸島に自生するゲッキツはアジアからオーストラリアの熱帯から亜熱帯に広く分布するとして、Murraya paniculataの学名が与えられ、3変種(var. omphalocarpa、var. ovatifoliolata、var. zollingeri)が区別されてきた。近年、形態学的観点から一部の変異群をM. exoticaとして区別すべきとの見解が提出されている(黄成就、1986)。var. omphalocarpaは台湾の属島蘭嶼(Botel Tobago)に特産するとしてナガミゲッキツの和名が田中長三郎により与えられたが、黄の見解ではM. paniculataに統合されている。その後の詳細な化学成分研究の結果、var. omphalocarpaはフィリピンに産する種と化学成分相にほとんど相違がないことが明らかになり、ナガミゲッキツはフィリピン以南の東南アジアに広く分布するものであることが明らかとなった。因みに南西諸島、台湾、中国南部に産するものは形態学的にM. exoticaとされるタイプであり、これまでに用いられてきた名称ゲッキツはこのタイプに継承される。化学的見地からナガミゲッキツとは異なるタイプのプレニルクマリンを含む(次図参照)ことで区別されることが明らかとなった。ゲッキツの化学分類の詳細についてはゲッキツ研究の概要および「木下武司 薬学雑誌 vol. 134 1265頁~1286頁 2014年」を参照。
2.フラボノイドとイソフラボノイド
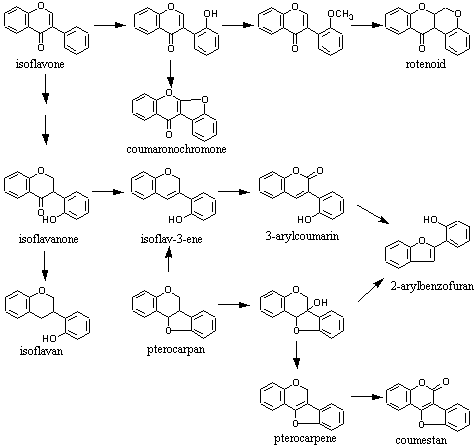
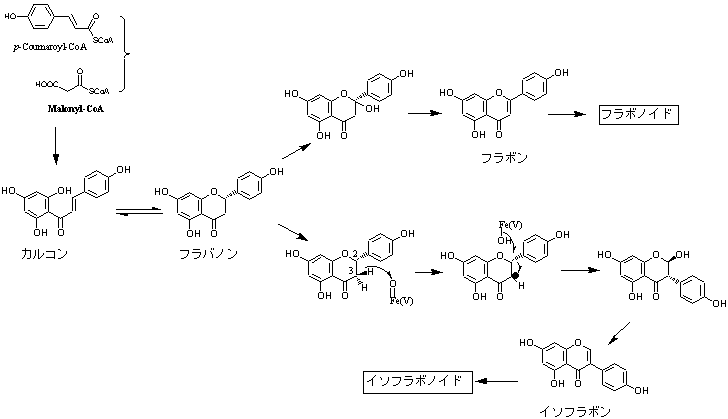
二次代謝産物にはある特定の生物種のみに出現するもの、限られた生物種に出現するもの、多くの生物種に出現するものなど、その分布の状態は多様である。主要な植物色素の一つであるフラボノイドは大半の植物に広く分布するが、構造的にその異性体であるイソフラボノイドはマメ科(Fabaceae)、特にソラマメ亜科(Papilinioideae)に集中し、同じマメ科でもジャケツイバラ亜科(Caesalpinioideae)、ネムノキ亜科(Mimosoideae)からきわめて限られた植物種から知られているにすぎない。マメ科以外ではアヤメ科アヤメ属など21科34属から報告されているが(J.B. Harborne Ed.,“The Flavonoids: Advances in Research since 1986”, Chapman & Hall,1996)、アヤメ属を除いてごく散発的に出現するのみでいずれも例外的存在として見なされている。マメ科に含まれるものは酸素ヘテロ環部が酸化、還元反応をうけて多様な骨格に分化しているのに対して、アヤメ属など非マメ科起源植物ではほとんどイソフラボンに限定されている(→イソフラボノイド各種を参照)。したがって、イソフラボノイドはソラマメ亜科を特徴づける特有の二次代謝産物群と考えて差し支えない。イソフラボノイドのうち、ロテノイドなど一部の成分群は限られた種に見られるなどの特徴があるが、それらを標識化合物とした化学分類学的研究はほとんどなされていない。フラボノイド、イソフラボノイドともに同じフラバノンを生合成前駆体とすることが知られている。次の図に示すように基本骨格の1,3-ジフェニルプロパノイドにおいてフェニル基が転移してイソフラボン骨格を生成するのであるが、この反応は生合成反応として特異な存在であるため、限られた植物種にのみ含まれ、二次代謝産物の化学進化の上でも興味深いものである。フラボノイド、イソフラボノイドの生合成については複合生合成経路を参照。
{kind=link}
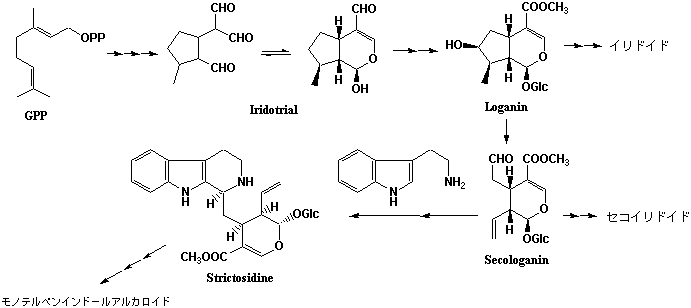
3.イリドイドとセコイリドイド
イソプレノイドの一種であるイリドイドも特定の植物群に集中して出現するものとして化学分類学の標識物質として用いられる。イリドイドの分布はミズキ上目(Cornanae)、ツツジ上目(Ericanae)、リンドウ上目(Gentiananae)、シソ上目(Lamianae)にほぼ限られており、特にセコイリドイドはリンドウ科(Gentianaceae)、マチン科(Loganiaceae)、キョウチクトウ科(Apocynaceae)、アカネ科(Rubiaceae)に集中し、リンドウ科ではスウェルチアマリン(Swertiamarin)やゲンチオピクロシド(Gentiopicroside)のように苦味物質として広く分布する。また、ロガニン(Loganin)から生成するセコイリドイドであるセコロガニン(Secologanin)はトリプタミンとともにモノテルペンインドールアルカロイド(→トリプトファン由来のアルカロイド)を構築し、マチン科、キョウチクトウ科、アカネ科ではこモノテルペンインドールアルカロイドを含む種と、アルカロイドを含まずイリドイド・セコイリドイド誘導体だけを含む種の両タイプが混在する。モノテルペンインドールアルカロイドに関しては、セコイリドイドに由来する部分がさらに様々な生合成反応を受けてさらに多様な骨格群に分化する。それぞれの植物種により特色ある骨格の成分が含まれるので化学分類学との相関は成立しやすい。モノテルペンインドールアルカロイドは様々な生物活性を有し医薬原料として有用なので、それらを標識物質として化学分類学が議論されがちであるが、本来はイリドイド・セコイリドイドを含めて討議されるべきである。植物分類学との関連からいえば、リンドウ科、マチン科、キョウチクトウ科、アカネ科はいずれも合弁花亜綱(Sympetalidae)に分類され、特にリンドウ科、マチン科、キョウチクトウ科はリンドウ目(Gentianales)に分類されている。リンドウ目にはこの他にガガイモ科Asclepiadaceae(APGではキョウチクトウ科に含められている)があるが、イリドイドは知られていない。 一方、アカネ科はアカネ目として別に分類されている。イリドイドを標識物質とした化学分類学は高度の分類学的知識が必要で議論される内容も難解であるが、イリドイドの生合成研究が詳細にわたって展開され経路がほぼ解明されていることが大きい。イリドイド、セコイリドイドの生合成はイソプレノイドの生合成(1)を参照。
4.ベンジルイソキノリンアルカロイド
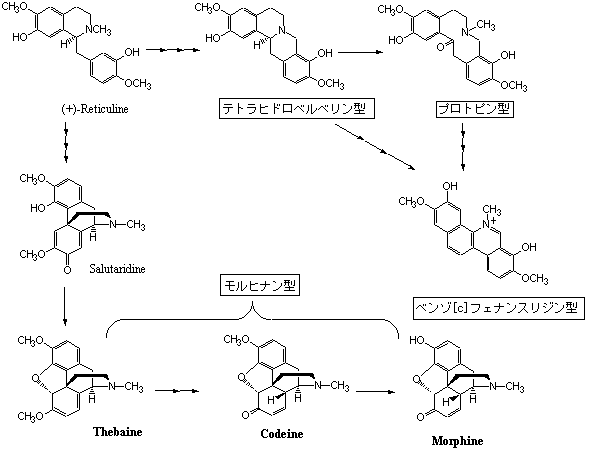
その他、生合成経路において同じ系列に属するもので、特定の生物種のみに出現するもの、限られた生物種に出現するもの、多くの生物種に出現するものがある。その具体的な例としてベンジルイソキノリンアルカロイドの分布を挙げて説明しよう。ベンジルイソキノリン系アルカロイドと称するものには多様な構造のものが知られているが、これらは全てレチクリン(Reticuline)と称する単純ベンジルイソキノリンアルカロイドから生合成される。その中で、レチクリンを含めて単純ベンジルイソキノリンアルカロイドは系統的には関係のないと思われる多くの植物群に含まれるが、これにC1ユニット(ベルベリンブリッジと称される)が導入されて生合成されるテトラヒドロベルベリン型アルカロイドではその分布は限られてくる。これまでのところ、離弁花亜綱(Archichlamiidae)に属する植物群からのみ報告され、キンポウゲ目(Ranunculales)に属するキンポウゲ科(Ranunculaceae)、ツヅラフジ科(Menispermaceae)、メギ科(Berberidaceae)の3科、同ケシ目(Papaverales)のケシ科(Papaveraceae)、同モクレン目(Magnoliales)のバンレイシ科(Anonaceae)、ムクロジ目(Sapindales)のミカン科(Rutaceae)から見いだされているに過ぎない。キンポウゲ目、ケシ目、モクレン目はモクレン下綱(Magnoliatae)に分類される系統的にまとまりのある植物群であるが、ムクロジ目はバラ下綱(Rosiatae)に属し前3目とは必ずしも系統的に近いとはいえない。一方、プロトピン型アルカロイドではケシ科のみに出現するが、ベンゾ[c]フェナンスリジン型ではミカン科とケシ科という系統的に遠い関係にある植物群に隔離分布する。ケシ科とミカン科ではベンゾ[c]フェナンスリジン骨格の置換様式が異なることが知られているので生合成経路が異なる可能性がある。ケシ科ではプロトピンを経てベンゾ[c]フェナンスリジンが生合成され、ミカン科ではプロトピンの生成を経ないで生合成される仮定すれば矛盾は解消される。本ページの冒頭で述べたように、全ての二次代謝産物に対して生合成経路が明らかになっている訳ではないので、化学分類学の適用には制限があることを念頭に入れるべきである。構造的にはフェナンスレン骨格とされるモルヒネ(Morphine)、コデイン(Codeine)、テバイン(Thebaine)などモルヒナンアルカロイドは生合成的にはレチクリンからサルタリジンを経て生合成されるベンジルイソキノリンアルカロイドの一種で、レチクリンからそれほど長くない生合成過程を経ているにすぎないが、ケシ科ケシ属の特定の数種(ケシPapaver somniferum、アツミゲシP. somniferum subsp. setigerum、ハカマオニゲシP. bracteatum)に見いだされるに過ぎない。ただし、光学的に鏡像体の関係にあるd-モルヒナンはツヅラフジ科オオツヅラフジやアオツヅラフジなどに見出される(シノメニンなど)。モルヒナン骨格を構築する生合成反応は酸化カップリングという、決して特殊な反応とは思えないが、テトラヒドロイソキノリンにおいてモルヒナンという複雑な骨格の形成を起こすことは自然界において化学的に特異な現象であることを示している。ベンジルイソキノリンアルカロイドの生合成はフェニルアラニン、チロシン由来のアルカロイドを参照。