アセチルCoAとマロニルCoAは生体内では一次代謝産物である脂肪酸の前駆体とされる。アセチルCoAとマロニルCoAはアシルキャリヤープロテイン(acyl carrier protein; ACP)に結合し、脱炭酸を伴う縮合が起きてジケチド体のアセトアセチル誘導体を生成する。この時、ケトン基の還元、脱水あるいはその結果生成する二重結合の還元も起きる。以上の反応が繰り返されて脂肪酸が合成されるのである(図1)。通例、脂肪酸鎖が偶数である(ステアリン酸:C18、パルミチン酸:C16)のはアセチルCoAにマロニルCoAに由来する炭素が2個単位で結合する生合成経路で生成するからである。一方、ケトンの還元を伴わずにC2の炭素鎖の延長が進行した場合、”ポリケチド(polyketide)”と称する非常に反応性の高い中間体を生成する。多くの場合、ポリケチドはアルドール型の縮合環化反応が起きて様々な芳香族化合物に変換される。
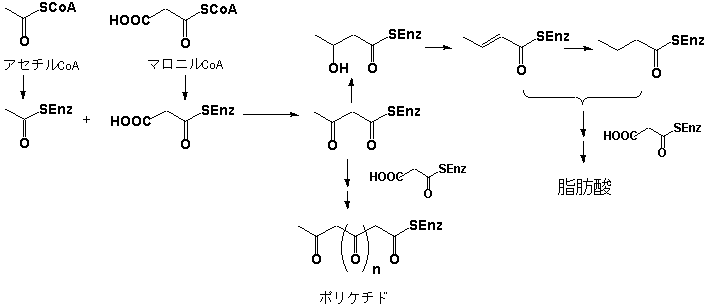
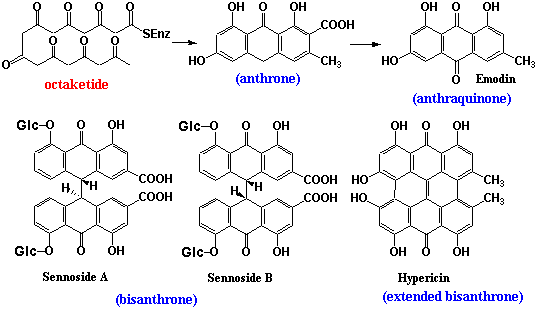
代表的なポリケチド由来の二次代謝物としてアントラキノン(anthraquinone)、アンスロン(anthrone)が挙げられるが、図2に示すようにアセチルCoAと7個のマロニルCoAから生成したオクタケチド(octaketide)に由来するものである。オクタケチドの閉環によりまず生成するのはアンスロンであり、脱炭酸、酸化を経てアントラキノンとしてもっとも広く分布するエモジン(Emodin)を生成する。アンスロンのメチレン部は反応性が高く、酸化カップリングによりしばしば二量化する。その代表的な物質としては緩下剤として繁用される生薬ダイオウ、センナの有効成分であるセンノサイド(センノシドともいう;Sennoside
A, B etc)や、オトギリソウ属(オトギリソウ、セイヨウオトギリソウ、コゴメバオトギリなど)に含まれるヒペリシン(Hypericin)などがある。ヒペリシンでは更に酸化カップリングが進行して完全に共役化しており、光増感物質としての性質を持つに至っている。タデ科ソバの茎葉にファゴピリンというヒペリシンに二分子のピペリジンが結合した特異なアルカロイドが含まれるが、やはり共役ビスアンスロン環に基づく光増感作用がある。アロエの瀉下作用成分にバルバロイン(Barbaloin)があるが、アンスロンのメチレン部に糖がC-C結合で結合した加水分解できない配糖体となっている。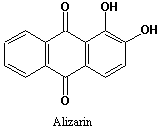 一方、アカネの根に含まれる色素アリザリン(Alizarin)は骨格としてはアントラキノンそのもの(右構造式)であるが、生合成的にはポリケチドではなく、シキミ酸-
一方、アカネの根に含まれる色素アリザリン(Alizarin)は骨格としてはアントラキノンそのもの(右構造式)であるが、生合成的にはポリケチドではなく、シキミ酸-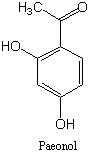 イソプレノイドの複合経路で精製するものである(→こちらを参照)。一般に、ポリケチド由来の二次代謝産物はその構造にポリケチド特有の特徴を残していることが多い。例えば、下記のアントラキノンでは酸素官能基は一つおきに配置しておりC2のアセテート単位のカルボニルの名残であり(→ポリケチド則)、同じアントラキノンでも非ポリケチド由来のアリザリンにはこのような特徴は見られない。その他、漢方生薬であるボタンピ(ボタンの根皮)に含まれるアセトフェノン誘導体ペオノール(左構造式;paeonol)も上記の特色が見られるのでポリケチド由来と推定することができる。
イソプレノイドの複合経路で精製するものである(→こちらを参照)。一般に、ポリケチド由来の二次代謝産物はその構造にポリケチド特有の特徴を残していることが多い。例えば、下記のアントラキノンでは酸素官能基は一つおきに配置しておりC2のアセテート単位のカルボニルの名残であり(→ポリケチド則)、同じアントラキノンでも非ポリケチド由来のアリザリンにはこのような特徴は見られない。その他、漢方生薬であるボタンピ(ボタンの根皮)に含まれるアセトフェノン誘導体ペオノール(左構造式;paeonol)も上記の特色が見られるのでポリケチド由来と推定することができる。
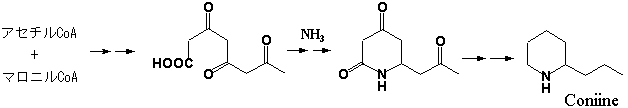
アルカロイドの中でアミノ酸を生合成前駆体としないものを”プソイドアルカロイド”と称するが、その中でポリケチドアミンと称するものがある。かつてシダ植物であるヒカゲノカズラ属(Lycopodium
sp.)由来のアルカロイドがポリケチド起源であるとする生.gif) 合成仮説があったが、実際にはリジン由来のものであった。これまでポリケチド由来の炭素骨格にアンモニア性窒素が取り込まれたことが実験的に証明されたのは極めて少ない。セリ科ドクニンジンの猛毒成分であるコニイン(Coniine)はそんな数少ないポリケチドアミンの一つである。本物質は未熟果実に多く含まれ、他に数種の類縁化合物が共存する。一方、ザクロ科ザクロの樹皮などに含まれる条虫駆除成分イソペレチエリン(Isopelletierine)は構造式こそコニインに酷似する(右構造式)が、リジンとジケチド(アセトアセチルCoA)による複合経路で生合成されるものである。
合成仮説があったが、実際にはリジン由来のものであった。これまでポリケチド由来の炭素骨格にアンモニア性窒素が取り込まれたことが実験的に証明されたのは極めて少ない。セリ科ドクニンジンの猛毒成分であるコニイン(Coniine)はそんな数少ないポリケチドアミンの一つである。本物質は未熟果実に多く含まれ、他に数種の類縁化合物が共存する。一方、ザクロ科ザクロの樹皮などに含まれる条虫駆除成分イソペレチエリン(Isopelletierine)は構造式こそコニインに酷似する(右構造式)が、リジンとジケチド(アセトアセチルCoA)による複合経路で生合成されるものである。